You better look twice or the importance of the not-so-cryptic species
The issue of the number of species on the planet always comes back to the headlines of the scientific journals. We have addressed this topic before in Mapping Ignorance focusing on a study 1 which estimated that even in theoretically well explored areas such as Europe we have not reached yet the “saturation plateau” of new species. That study would be yet another example of the perspective, arguably started with Erwin’s works on tropical beetles2, stating that there may be a good bunch of millions of species yet to be described, and that our degree of knowledge of them is ridiculously low. In contrast, we have also read in the past months other papers concluding that actually most of the species are probably already catalogued by science34. Which one is right? At this point, we obviously do not know the answer to the 300-year-old question, and in fact, what makes this issue not just old, but also challenging and frustrating at the same time is that we still do not even know the order of magnitude of our own ignorance.
Given that a direct count is not possible (that is why we cannot answer the question in the first place), all we can do are Fermi approximations, educated guesses with our available data. The problem, of course, is that the quality of those data may affect dramatically the resulting confidence interval. Many estimations are calibrated with groups that should be, at this point, “well known”, meaning that the real number of species of a certain lineage may not be very far from the current taxonomical knowledge. Arguably, certain groups of vertebrates on sufficiently explored areas would fulfill this requirement, and some estimates are actually extrapolations based on these constrains. The dilemma, however, remains: how confident can we be about “well known groups”?
Four researchers from several Australian institutions recently arqued5 that these assumptions may be wrong because they ignore the “elephant in the room”: cryptic species. Although the definitions of this term vary, two species are considered cryptic if they are difficult or impossible to set apart by “traditional means”, usually meaning morphological analysis. If cryptic species are rampant, most of the times that a biologist identifies an organism it will be ascribed to an already described taxon even if it belongs to an “unknown” species, and this could mean that our collections in museums and herbaria are paradoxically packed with animals and plants unknown to science.

Although this perspective is not new6, Adams et al. claim that cryptic species and their impact are not seriously accounted in current diversity estimations. To reflect this idea, they show the result of their revision of Galaxias olidus, a freshwater fish from Southeastern Australia whose taxonomy should be considered “largely finalized”. Their integrative approach included the morphological study of more than 3000 specimens, a comprehensive isozyme assessment of over 800 fishes and DNA barcodes of 557 of these animals. This extensive work was rewarded with an amazing answer: Galaxias olidus is not such a well-known fish species, but a hyper-cryptic complex of 15. It is perhaps tempting to think that this conclusion is just the result of applying molecular phylogenetics to a group of organisms that otherwise would have remained invisible to old-school taxonomy, but as the authors themselves remark, all these 15 species are indeed morphologically diagnosable. This, of course, makes us to reconsider when a species should be considered truly cryptic: it is not rare to find publications that conclude after a phylogenetic analysis that a number of unnoticed lineages are new species that were not noticed by morphological studies because they were cryptic, but what the Galaxias olidus study suggests is that morphological reevaluations are critical for a diversity assessment. How frequently new species are just considered “cryptic” because a morphological revision is lacking? I suspect this happens very often.
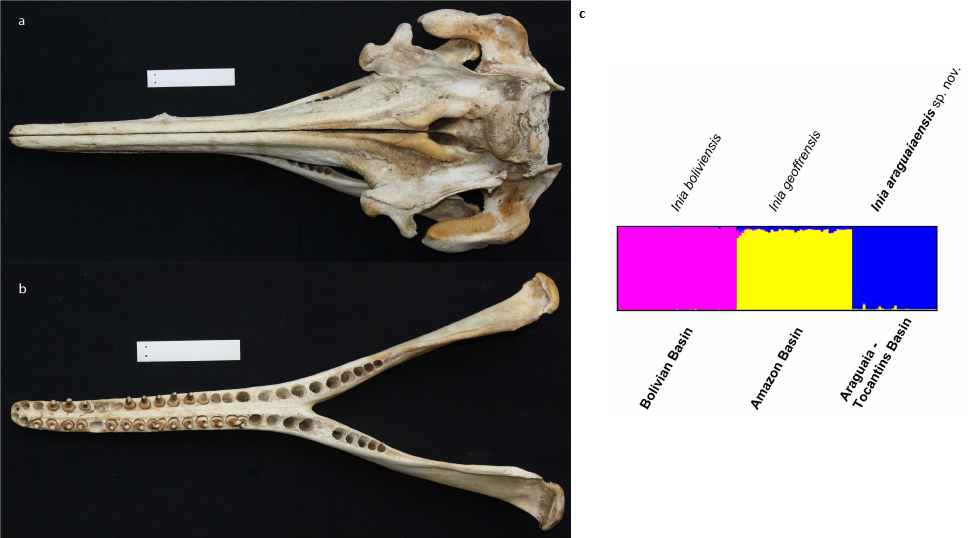
Recently, a new species of river dolphin (Inia araguaiaensis) was described in the Araguaia basin, Brazil7. Cetaceans are conspicuous mammals that attract a lot of attention, animals that we could also consider “well known”. It is remarkable that already in the 21st Century this dolphin was still unrecognized as distinct from the Amazonian Inia geoffrensis and I. boliviensis, and yet this discovery should appear on textbooks as an example of a neat and beautiful discovery of a new species: the araguaian dolphin fulfills the biological species concept (its geographic distribution and microsatellite spectrum suggests reproductive isolation), the phylogenetic species concept (according to the cytochrome b sequence) and the morphological species concept (it shows less teeth than the Amazonian species and bears different cranial morphometric indices). This dolphin was not cryptic at all, there was no technical impediment for its discovery even in the pre-molecular era, it just did not receive enough attention. If this can happen with a mammal, how not to be thrilled about truly inconspicuous groups?
There is a growing evidence provided by integrative studies showing good correlations of molecular phylogenetics and modern morphological revisions, even in minute organisms with a limited morphospace such as proturans8 or liverworts9. The point is that even if cryptic species are a real challenge for biodiversity assessment, it should not be taken for granted that morphological studies have already reached its limit. In words of Adams et al.5:
[…]the ultimate and crucial taxonomic outcome of diagnosing candidate species requires considerable intensive effort in two key traditional and labour-intensive components of taxonomic endeavour: targeted collecting of vouchers, and detailed morphological analyses i.e., there are no shortcuts to reach the final taxonomic outcome.
References
- Fontaine, B. et al. New species in the Old World: Europe as a frontier in biodiversity exploration, a test bed for 21st century taxonomy. PLoS One7, e36881 (2012). ↩
- Erwin, T. Tropical forests: their richness in Coleoptera and other arthropod species. Coleopt. Bull. (1982). at
- Costello, M. J., Wilson, S. & Houlding, B. Predicting total global species richness using rates of species description and estimates of taxonomic effort. Syst. Biol.61, 871–83 (2012). ↩
- Appeltans, W. et al. The magnitude of global marine species diversity. Curr. Biol.22, 2189–202 (2012) ↩
- Adams M., Raadik T.A., Burridge C.P. & Georges A. Global Biodiversity Assessment and Hyper-Cryptic Species Complexes: More Than One Species of Elephant in the Room?, Systematic Biology, DOI: 10.1093/sysbio/syu017 ↩
- Bickford, D. et al. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol.22, 148–155 (2007) ↩
- Hrbek, T. et al. A New Species of River Dolphin from Brazil or: How Little Do We Know Our Biodiversity. PLoS One 9, e83623 (2014) ↩
- Resch, M. C. et al. Where taxonomy based on subtle morphological differences is perfectly mirrored by huge genetic distances: DNA barcoding in protura (hexapoda). PLoS One9, e90653 (2014) ↩
- Renner, M. A. M. et al. Integrative taxonomy resolves the cryptic and pseudo-cryptic Radula buccinifera complex (Porellales, Jungermanniopsida), including two reinstated and five new species. PhytoKeys 1–113 (2013). doi:10.3897/phytokeys.27.5523 ↩
4 comments
[…] It turns out that the blue group corresponds precisely to the populations under the name of Narcissus bujei, and its distinction from the rest of the forms of the section is strongly supported. According to the authors, however, the […]
[…] You better look twice or the importance of the not-so-cryptic species […]
[…] different species can be hard to tell apart. An extreme case involves cryptic species. These are separate species that are very similar outwardly, yet are true species that never […]
[…] est difficile, parfois, de distinguer des espèces différentes. Cas extrême : les espèces cryptiques. Extérieurement, elles sont semblables, mais elles se révèlent être des espèces spécifiques […]