Cultural traditions in orcas
Cultural traditions in orcas

Orcas or killer whales (Orcinus orca) are widely distributed and feed on a large array of prey —fish, seals and sometimes whales—. These powerful predators are highly adaptable, cooperative and live in stable matriarchal social groups of related animals. Each prey species they feed on requires special hunting skills and there have been observational reports of astonishing foraging tactics. A new study suggests they learn by imitation.
A few examples of these skills include the intentional stranding or beaching of killer whales in order to catch seal pups in the Crozet Archipielago (southern Indian Ocean)1 and in Punta Norte, Argentina 2. Studies have shown that the orcas can time their incursions onto land to coincide with the tides, so they run less risk of becoming beached permanently. Apprenticeship appears to play a very important part in this behaviour, since mothers and other adult pod members practice intentional stranding with calves. Argentinean orcas have been seen nudging youngsters onto the shore, encouraging them to try the tactic, often coming up alongside to demonstrate. Other example of cooperative hunting behaviour takes place in the Antartic waters, where orcas use cooperative wave-washing behavior 3 —producing a large number of waves— in order to wash up seals from the ice floes. Pod elders seem to teach younger animals —in the Antartic Peninsula young orcas are often present during the hunt and adults sometimes put living seals back on the ice after catching them, so that the young may have another try. In Northern Norway researchers have observed a different cooperative behaviour called the ‘carousel feeding’ technique 4. Killer whales cooperative herd herring into a tight ball close to the surface, swim around and under the fish emitting large bubbles and stunning their prey by slapping the edge of the school with the underside of their flukes. Once the fish is stunned, they eat them one by one. In New Zeland waters killer whales were observed benthic foraging on stingrays in shallow water using the bottom as a physical barrier to trap them and preventing them from escaping 5. Lastly, in the area of the Strait of Gibraltar, killer whales chase bluefin tuna of small to medium size for up to 30 minutes at a relatively high sustained speed (pushing tuna beyond their aerobic limits to exhaust them), until they captured them —named the ‘endurance-exhaustion’ technique 6.
When one reads about this array of foraging tactics, a question that comes to mind is how are all these behaviours learned? Are they cultural traditions, in the same way as the potato washing of Japanese macaques or the nut cracking of chimpanzees? Is imitation the main social learning mechanism involved or these ‘cultural traditions’ can be explained by other less complex learning processes?
Cetaceans are likely candidates to display imitative learning since they have evolved in socioecological backgrounds that have selected for large brains, social complexity, flexible communication systems and coordinated predatory tactics. Some authors support the view that the large brain of cetaceans evolved to support complex cognitive abilities —impressive cultural learning of dialects, foraging sites and hunting strategies, self-recognition 7.
A team of researchers from the Universidad Complutense de Madrid (Grupo de Estudio del Comportamiento Animal y Humano) and the Max-Planck Institute for Evolutionary Anthropology have recently published a paper 8 in which they carried out an experimental study with the aim of discovering whether killer whales are able to imitate on command the actions performed by a conspecific demonstrator. If the behaviour copied by the ‘observer’ from the ‘demonstrator’ is novel, does not exist in the observer’s repertoire, we can affirm that ‘complex imitation’ or ‘imitative learning’ has taken place, entailing more complex cognitive processes since it requires a new motor schema in its pre-existing repertoire.

In this study the method ‘do-as-other-does’ was used to test killer whales’ capacity to imitate both familiar and novel actions (15 familiar and 4 novel behaviours). Researchers tested 3 killer whales housed at an aquarium in France —Val, a 13-year-old male; Inouk, a 10-year-old male; Wikie, a 8-year-old female—, all borned in captivity, mother-reared and trained for a variety of exercises with standard operant conditioning procedures and fish/tactile positive reinforcement. They conducted 48 experimental sessions (see experimental set up in figure 1) that were videotaped by two video cameras. Two trainers were needed, one to give the signal to the demonstrator (TD) and another to give the copy command to the subject (TS). The study comprised three phases. In Phase 1 subjects were trained to respond to the gesture-based command ‘copy’ (‘Do that’) given by the trainer. In Phase 2 they tested the subjects’ responses to the trainer’s copy command when the demonstrator performed familiar actions —all familiar behaviours were not part of the killer whales’ natural repertoire but the result of the trained series of actions that the orcas were requested to perform during the aquarium shows (see table 1 of behaviours tested in each phase).
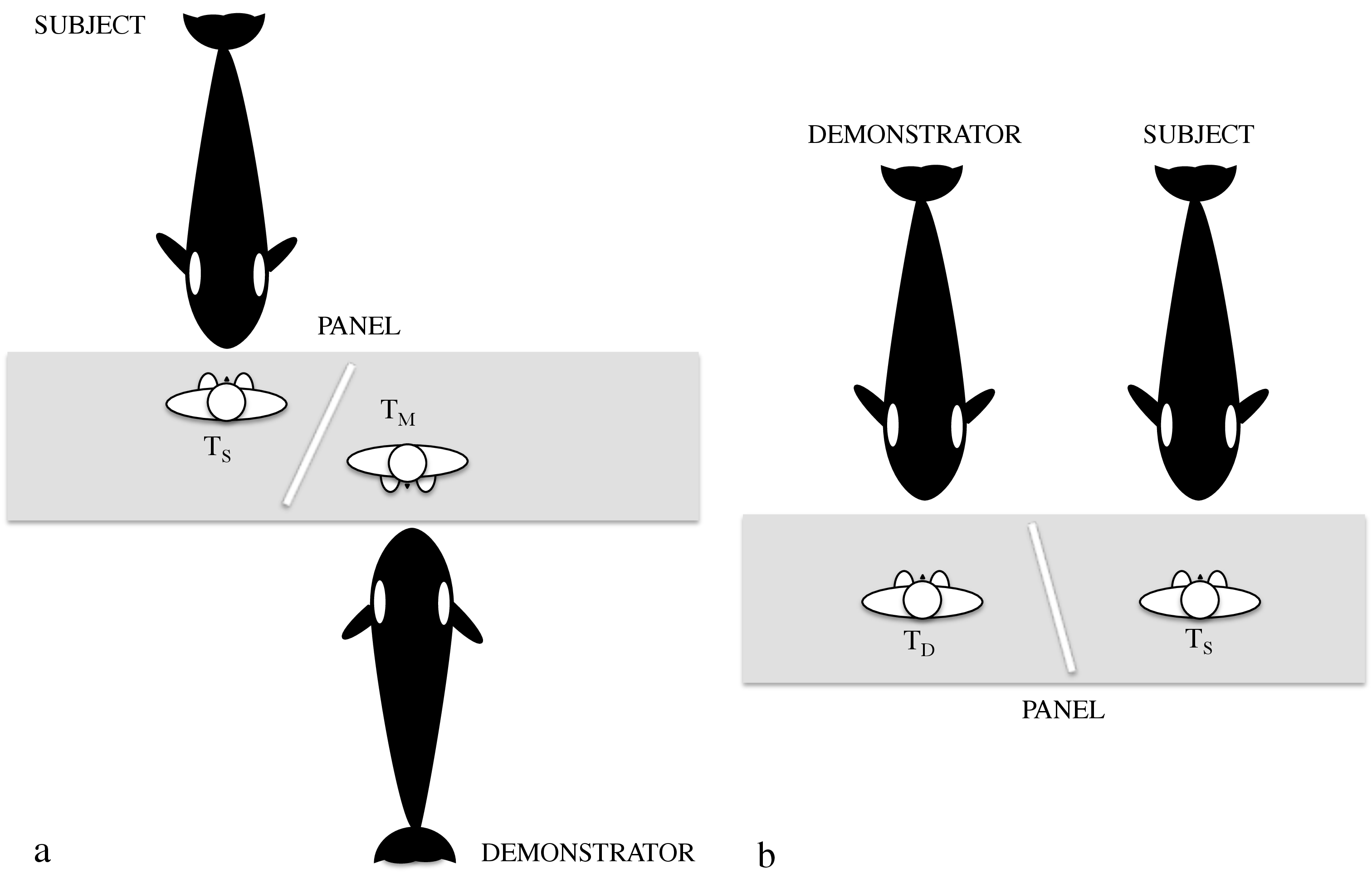
In Phase 3 the subjects were tested with novel behaviours, that is, behaviours that were unknown to the subject. In all testing sessions, both trainers were positioned at different sides of a white wooden panel, placed in a way that the subject and the demonstrator could see each other and their trainer, but could not see the other trainer’s signal.
Coding of videos was carried out by experimenters who had not been present during the training sessions. Coder 1 coded the videos several hours after the testing sessions, and coder 2 several months after the study had been completed (the second coder could see the subject’s actions but could not see the trainer’s signal). Interobserver reliability was excellent. For assessing familiar behaviour, researchers used the level of matching accuracy of each attempt with a three-point scale: (1) for full reproduction, they gave a 1; (2) for partial reproduction (some elements of the modelled action were missing), they gave a 0.5; (3) for failed reproduction, they gave a 0 (the action performed was completely unrelated to the one performed by the demonstrator). For novel behaviours they added a fourth category, (4) ‘almost full reproduction’ (0.75), in order to identify any minor differences in the behaviour matching.
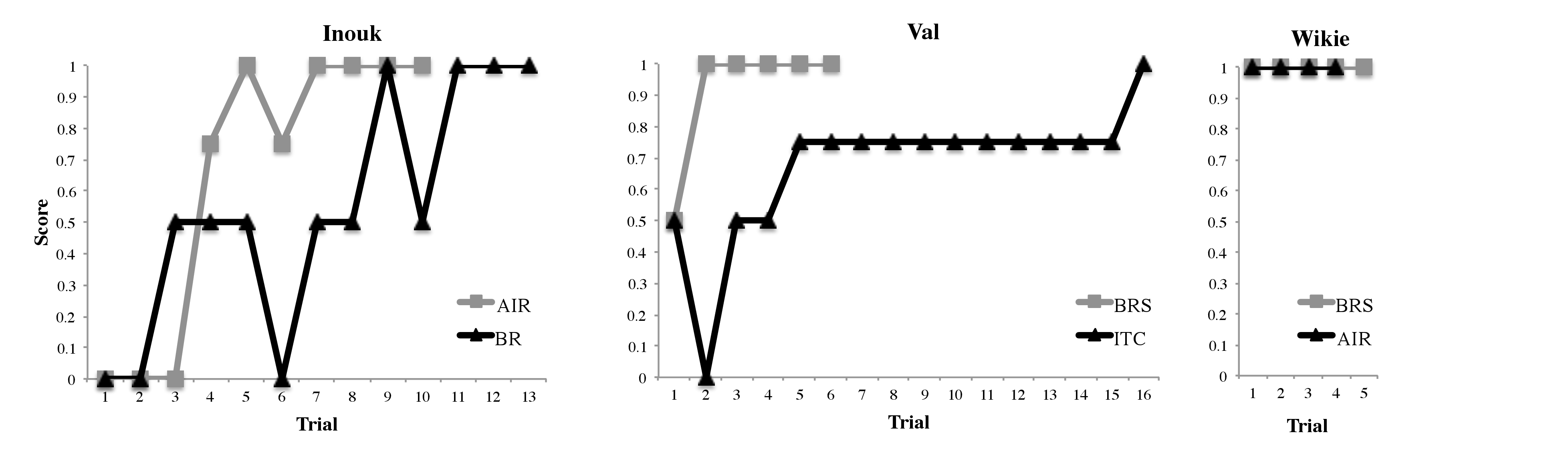
The results showed that all three orcas learned the copy command signal ‘Do that’ rather fast, in 20 trials on average; they copied 100% of the demonstrator’s familiar and novel actions; they accomplished full matches in the first attempt for 8-13 familiar behaviours (out of 15) and for the 2 novel behaviours (out of 2) in one subject; all three took no longer than 8 trials to accurately copy any familiar behaviour, and no longer than 16 trials to copy any novel behaviour. Figure 3 shows the subject’s performance in each trial for each novel actions analysed. One can see that on the few trials in which the killer whales made mistakes, most of them reproduced the major components of the demonstrator’s action.
As we have seen in the examples at the beginning of this article, there are cases of teaching in the wild in which mother killer whales and other adults in the pod ‘instructed’ calves in the intentional stranding hunting technique. In this study the authors report an anecdote that may illustrate cetaceans natural inclination to copy others’ actions. The 2-month-old calf of the female Wikie, who was always by her side, spontaneously performed a half of the untrained behaviour ‘Airplane’ and an accurate match of two familiar behaviours, ‘Fluke wave’ and ‘Song’, produced by her (there are five movies available as supplementary material in the paper online).
In summary, this study adds new experimental data on cetacean imitation, adding support for action imitation in the delphinids, since experimental evidence for vocal imitation and action imitation had only been demonstrated before in the bottlenose dolphin. I consider these results very important since most studies on imitation had been carried out in birds and great apes. In the near future there will probably appear new reviews of imitation in chimpanzees, killer whales and dolphins thus nourishing the animal culture debate.
References
- Guinet C, Bouvier J (1995) Development of intentional stranding hunting techniques in killer whale (Orcinus orca) calves at Crozet Archipielado. Can J Zool 73:27-33 ↩
- Lopez JC, Lopez D (1985) Killer whales in Patagonia and their behaviour of intentional stranding while hunting nearshore. J Mammal 66:181-183 ↩
- Pitman RL, Durban JW (2012) Cooperative hunting behaviour, prey selectivity and prey handling by pack ice killer whales (Orcinus orca), type B, in Antartic Peninsula waters. Mar Mammal Sci 28:16-36. DOI: 10.1111/j.1748-7692.2010.00453.x ↩
- Similä T, Ugarte F (1993) Surface and underwater observation of cooperatively feeding killer whales in Northern Norway. Can J Zool 71:1494-1499 ↩
- Visser IN (1999) Benthic foraging on stingrays by killer whales (Orcinus orca) in New Zeland waters. Mar Mammal Sci 15:220-227 ↩
- Guinet C, Domenici P, de Stephanis R, Barrett-Lennard LG, Ford JKB, Verborgh P (2007) Killer whale predation on bluefin tuna: exploring the hypothesis of the endurance-exhaustion technique. Mar Ecol Prog Ser 347:111-119 ↩
- Marino, L, Connor, RC, Fordyce, RE, Herman, LM, Hof PR et al. (2007) Cetaceans have complex brains for complex cognition. PloS Biol 5 (966-972): e139. DOI: 10.1371/journal.pbio.0050139 ↩
- Abramson, JZ., Hernández-Lloreda, MV, Call, J., Colmenares, F. (2013). Experimental evidence of action imitation in killer whales (Orcinus orca). Animal Cognition 16, 1 (11-22). DOI: 10.1007/s10071-012-0546-2 ↩
4 comments
[…] Humans aren’t the only animal species to share traditions. When orcas happen upon a useful way of doing things, they pass it along. […]
[…] bonds between orca are fascinating: each pod has a unique dialect, distinct food preferences, and cultural […]
[…] aux générations suivantes. Différentes populations locales ont développé leurs propres techniques de chasse tel […]
[…] passed down to subsequent generations. Different local populations have developed their own favored hunting techniques such […]