Trident whiskers
Trident whiskers

Whiskers or vibrissae are a type of mammalian hair characterized, anatomically, by their larger, thicker and stiffer morphology, large and well-innervated hair follicle and by having a definite projection in the somatosensory cortex of the brain, forming the so-called “barrels”. The ‘barrels’ of the barrel field are regions within cortical layer IV that are visibly darker when stained to reveal the presence of cytochrome c oxidase, and are separated from each other by lighter areas called septa. A rat has around 300 whiskers.
Whiskers typically grow in groups in different locations, usually conserved throughout evolution. In the head, many land animals have supraorbital (above the eyes), genal (of the cheeks), and mystacial (where a moustache would be), as well as mandibular (of the jaw) vibrissae under the snout.
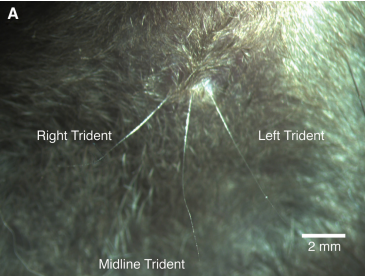
There is also an under-investigated submandibular «whisker trident» on the chin. It consists of three whiskers: an unpaired midline whisker and a pair of mirror-symmetric slightly shorter whiskers flanking it. All three whiskers point to the ground and are curved backwards. The trident whiskers have many peculiarities 1:
-
Their unexposed position on the animal’s chin. They do not touch anything over long stretches of time.
-
Their selective engagement in ground contact during head-down running while the initial exploration of novel environments. Remarkably, habituation was very fast and deflection angles decrease with increasing ground velocity and after a second exposure to the same environment.
-
The unpaired arrangement of the midline trident whisker, the only unpaired whisker of the vibrissal system.
-
The elongated putative trident-midline barrel. The trident-midline barrel is the longest and largest whisker barrel, suggesting a great functional significance.
-
The unique asymmetrical cortical representation of the midline trident. Thé and coworkers found one putative trident barrel in the left somatosensory cortex and two barrels in the right somatosensory cortex. Thus, there is a striking hemispheric asymmetry. In addition, most whisker barrels in the rat cortex are sharply delineated, but this was note the case for the putative trident barrels.
-
Their biomechanical ground sensing capacities. Set angles were ground-speed dependent.
-
The slower kinetics of cortical neurons to trident whisker deflections as compared to deflections of facial whiskers.
Cortical postsynaptic air-puff responses in the trident representation confirm the right hemispheric bias in the representation of the midline trident and show much less temporal precision than facial whisker responses. Trident and facial whiskers do not only differ in their morphology, behavioral use, and biomechanics but also show remarkable differences in the response dynamics. Trident whiskers do not provide as much high-resolution information about object contacts as facial whiskers. Instead, these observations suggest an idiothetic function: trident whiskers obtain continuous measurements about egomotion from ground contacts.
Egomotion information is the information regarding the motion of the subject in space and such information is of particular value to nocturnal animals, which navigate in the dark, and thus need to estimate their location in space. The midline position offers unique advantages in sensing heading direction in a laterally symmetric manner. Thé’s research team suggested that changes in trident deflection angle with velocity suggest that trident whiskers could be read out as a tactile speedometer.
A recent study by the same group has studied the cortical representation of trident whiskers and E-row whiskers in barrel cortex 2. E-row whiskers are the lowermost row of macrovibrissae and they are also often in contact with the ground during locomotion and thus suitable for monitoring locomotion behavior. The putative trident barrel columns areas project to the vestibular nuclei, as well as to the posterior parietal cortex, an area thought to be involved in pathway integration. These projections to the spinal vestibular nucleus are unique to trident cortex and do not extend to the nearby facial or paw somatosensory cortices.
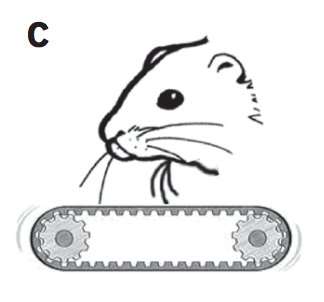
The authors wanted to analyzed if cells from anesthetized animals respond to ground displacement in the cortical trident barrel columns. To check this, they run a little treadmill under rat’s chin to look at movement representation in trident whisker defection. Second, they wanted to analyze whether egomotion-related responses were seen in awake, freely behaving, foraging rats. They assessed whether the firing of units in the trident, E-row barrels and the dysgranular zone (DGZ) were modulated by speed, acceleration and turns. The DGZ is a part of the somatosensory cortex between the forelimb and macrovibrissae representations and is not linked to cutaneous sensations and thought to be involved in proprioception.
Neurons identified in trident cortex of anesthetized animals showed sustained velocity-sensitive reactions to ground motion. While effects of trident and E-row cortex microstimulation on locomotion speed were common and highly significant, the changes in locomotion speed were typically minor. Given the intactness of other sources of egomotion information, such as vestibular inputs, the presence of an effect is meaningful. It implies that speed estimation is a multisensory operation, whose outcome directly affects the locomotion program. In freely moving animals, about two-thirds of the units in the trident and E-row whisker cortices were tuned to locomotion speed, a larger fraction of speed-tuned cells than in the somatosensory DGZ. Similarly, more units were tuned to acceleration and showed sensitivity to turning in trident and E-row whisker cortices than in the DGZ. Microstimulation in locomoting animals evoked small but significant speed changes (larger in the trident and E-row whisker representations than in the DGZ). Thus, activity in trident and E-row cortices represents egomotion information and influences locomotion behavior.
Information related to egomotion is found in many brain areas including V1 (primary visual cortex) even in the absence of optic stimuli. The brain can therefore calculate mismatch between the predicted speed, which is derived from the motor plan, and the perceived speed, which is derived from diverse sensory flow information.
References
- Thé L, Wallace ML, Chen CH, Chorev E, Brecht M (2013) Structure, function, and cortical representation of the rat submandibular whisker trident. J Neurosci 33(11): 4815-4824. ↩
- Chorev E, Ferrer PP, Brecht M (2016) Representation of egomotion in rat’s trident and E-row whisker cortices. Nat Neurosci. 2016 Aug 15. doi: 10.1038/nn.4363. ↩
1 comment
[…] Las ratas tienen “bigotes”, pero pocos saben que tienen una barba de tres pelos, como la de la canción infantil. Sin embargo, este tridente piloso tiene una utilidad increíble para la rata. JR Alonso en Trident whiskers. […]