Desert scavengers, amazing navigators
Desert scavengers, amazing navigators

Cataglyphis, the desert ant, is a genus in the subamily Formicinae. The most famous species is C. bicolor, the Sahara Desert ant. There, the midday sun is so hot that even the permanent residents, sand lizards, insects and a few birds, have to take shelter. But this is when, for not much than an hour, it is safe for Cataglyphis to come out of their underground nests and scavenger since all its predators are in hiding from the sun. During that time, they can sustain body temperatures well above 50 ºC with surface temperatures of up to 70 ºC and they run on hot sand to find insects’ and other arthropods’ corpses that succumbed of heat exhaustion, their food source.
Sahara ants live in terrains where there are no bushes or clumps of grass, odors quickly vanish and tracks are covered by wind-blown sand in seconds. Thus, there are almost no identifiable features in the environment but they need to return to their nest as fast as possible to avoid becoming a victim of the prevailing heat and dessication stress. Each ant travel thousand of times their own body length, dashes about in zigzag patterns, but as soon as one is lucky enough to find a tiny insect body, it has to get it back to the nest fast before it itself dies. The nest entrance is inconspicuous, only a 2 cm hole in the desert floor and the ant does not retrace the zig-zagging path of its outward journey; even if a scent-trail made this possible, such a route would be a waste of time and an increased risk. Instead, it runs in a straight line directly back to its nest-hole. It is amazing that the 0,1 mg brain housed in the head of this 10 mg ant accomplish such feats of navigation.
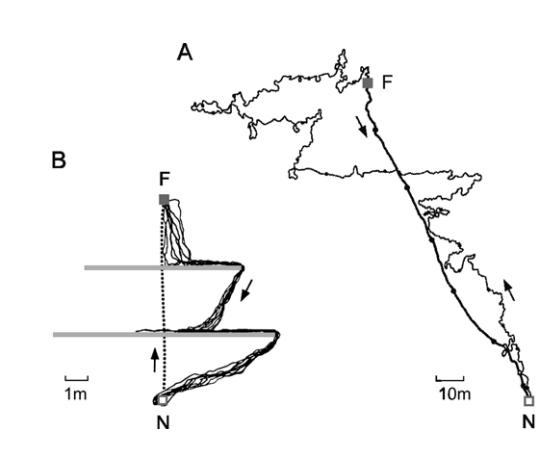
Cataglyphis continually updates spatial information about places and routes. The ants exhibit considerable flexibility in coupling and decoupling different navigation modules and referential systems to and from each other 1. Cataglyphis ants are used to study path integration, a navigation feat in which angles steered and distances travelled are combined to provide a direct link to the point of origin, a so-called home vector. In the path integration, they use two strategies: «stride integration» and «optic flow». Stride integration accounts for stride number and the respective stride length and is like wearing a fitness tracker —an odometer— to count steps. Optic flow refers to image motion experienced during travel and is akin to how we estimate distance and direction out the window when riding in a car. Ants are visual walkers, so stride integration and optic flow integration are difficult to tease apart from each other experimentally.
However, Sarah Pfeffer and Matthias Wittlinger, from University Ulm have devised an interesting approach using a particular behavior of these insects: Cataglyphis bicolor ants carry their nestmates to new nests in a «pupal» posture. This behavior is common in these ants because of their polydomous nest structure, where the main nest with the single queen is surrounded by satellite nests. Experience foragers that walk a set distance from the nest are able to return without issue because they count the steps it took to get to that point. In the first experiment, the researchers connected two nest entrances of neighboring nests with a walking channel 2. The carrier ant can count its steps, but the passenger ant can only watch as they travel, in an inverse fashion (legs up). Ants transported by nest mates, and thus without stride integration, are capable of measuring travel distance exclusively by the use of optic flow cues. When separated, the passenger ant can find its way back to the nest because that visual information tells it where to go. These experiments show that the exclusive use of optic flow cues was enough for the ants to measure travel distances. Therefore, active locomotion is not a prerequisite for distance estimation or path integration, provided that the transport happens in a naturally occurring behavior.
In a second set of experiments, the researchers blindfolded the ventral part of the compound eyes of a separate test group with bright yellow paint to see whether they could find their way back to the colony. It has already been shown that ventral eye caps do not interfere with proper homing behavior in walking ants and thus ventral blindfolding does not affect correct distance estimation. The ants of the blindfolded test group did not obtain any distance information in the outbound journey because visual input was eliminated (covered eyes) and the stride integrator was also not effective (carried mode). The blindfolded ants did not head for the home nest; their search was concentrated around the releasing point and they get lost.
There is ample evidence that the stride integrator can function without visual input. Ants walking in complete darkness, or with the ventral halves of their eyes covered, are still able to evaluate homing distance correctly. Although the interior worker did not know how many steps away their home was, they were able to rely on sight to return, which is a surprise because vision was only thought to play a supporting role in navigation 3. However, although the results show that two separate and independent systems of distance measurement exist and operate redundantly, the information gained from the optic flowmeter cannot be transferred to the stride integrator and they do not fill in for one another. Saharan habitat is so tough that the presumably the ants evolved two separate mechanism to cope.
References
- Wehner R (2003) Desert ant navigation: how miniature brains solve complex tasks. J. Comp. Physiol 189: 579-588. ↩
- Pfeffer SE, Wittlinger M (2016) Optic flow odometry operates independently of stride integration in carried ants. Science 353(6304): 1155-1157. doi: 10.1126/science.aaf9754 ↩
- Boddy J (2016) Video: ‘Blindfolded’ ants reveal clues to insect navigation. Science ↩
1 comment
[…] Las hormigas del Sáhara viven en zonas donde no hay ni matorrales ni siquiera hierba, donde los olores desaparecen rápidamente y las huellas las barre el viento en segundos. Y sin embargo, son capaces de orientarse con una eficiencia […]