The diversity conundrum: Why do oceans shelter fewer species than land?
The diversity conundrum: Why do oceans shelter fewer species than land?
Authors: Guillem Chust, ; Xabier Irigoien, ; and Naiara Rodríguez-Ezpeleta, Head of Molecular Ecology and Biotechnology at AZTI, Marine Research / Basque Research and Technology Alliance (BRTA)

Most nowadays existing animal groups originated in the sea after the Cambrian explosion, 540 million years ago, before there was life on land. In that time, the great diversity of life in the sea sheltered trilobites, stem-group arthropods such as the large predator Anomalocaris, crustaceans, sponges, early chordates, worms, and squid-like creatures. The beginning of extensive land colonization by plants was millions of years later, during the Devonian (~390 Ma), which was facilitated through interactions with symbiotic fungi. Long after this transition, the coevolution between flowering plants with insects spurred the vast diversification of both groups.
Is the dominance of terrestrial diversity an historical phenomenon or does it rather obey to general ecological theories? Is the Ocean a “deathtrap” favouring species extinction as suggested by Miller and Wiens in 2017? One of the well-known rules in ecology establishes that the number of species found in an area increases with the size of the area. As oceans cover 71% of the Earth’s surface and have harboured life for much longer than land, one would expect that oceans hold a greater number of extant species than land. However, less than 15% of multicellular eukaryotic species currently described are marine, 80% live on land and 5% in freshwater. This diversity conundrum was pointed out by Robert May in 1994.
Although widely recognized, explanations for the diversity conundrum have often been limited to particular species groups or focused on testing specific hypotheses, such as how specific terrestrial environments permitted high speciation. In our paper 1, we get deeper into the diversity conundrum by focusing on the limitations of marine pelagic diversity compared to land and freshwater diversity.
Biodiversity hotspots and highly diversified groups
There are 2.16 million described species in our planet. One challenge underpinning different biodiversity patterns across land and ocean is that undescribed and cryptic species may be more abundant in marine environments. The difficulty of exploring remote and deep seas, and the estimated high number of species still undescribed could alter the estimated number of species in each realm. There are currently ca. 247,447 known valid marine species living in the oceans, which continue to be discovered steadily at a current average of 2,332 new species per year, i.e. 0.95% over the total marine species richness. The rate of terrestrial species description is about 16,000 species per year, i.e. 1.02% over the total terrestrial species richness, a percentage similar to that of marine species. Therefore, it is very unlikely that sampling bias would be the sole explanation for the observed imbalance of life diversity comparing oceanic and terrestrial biomes.

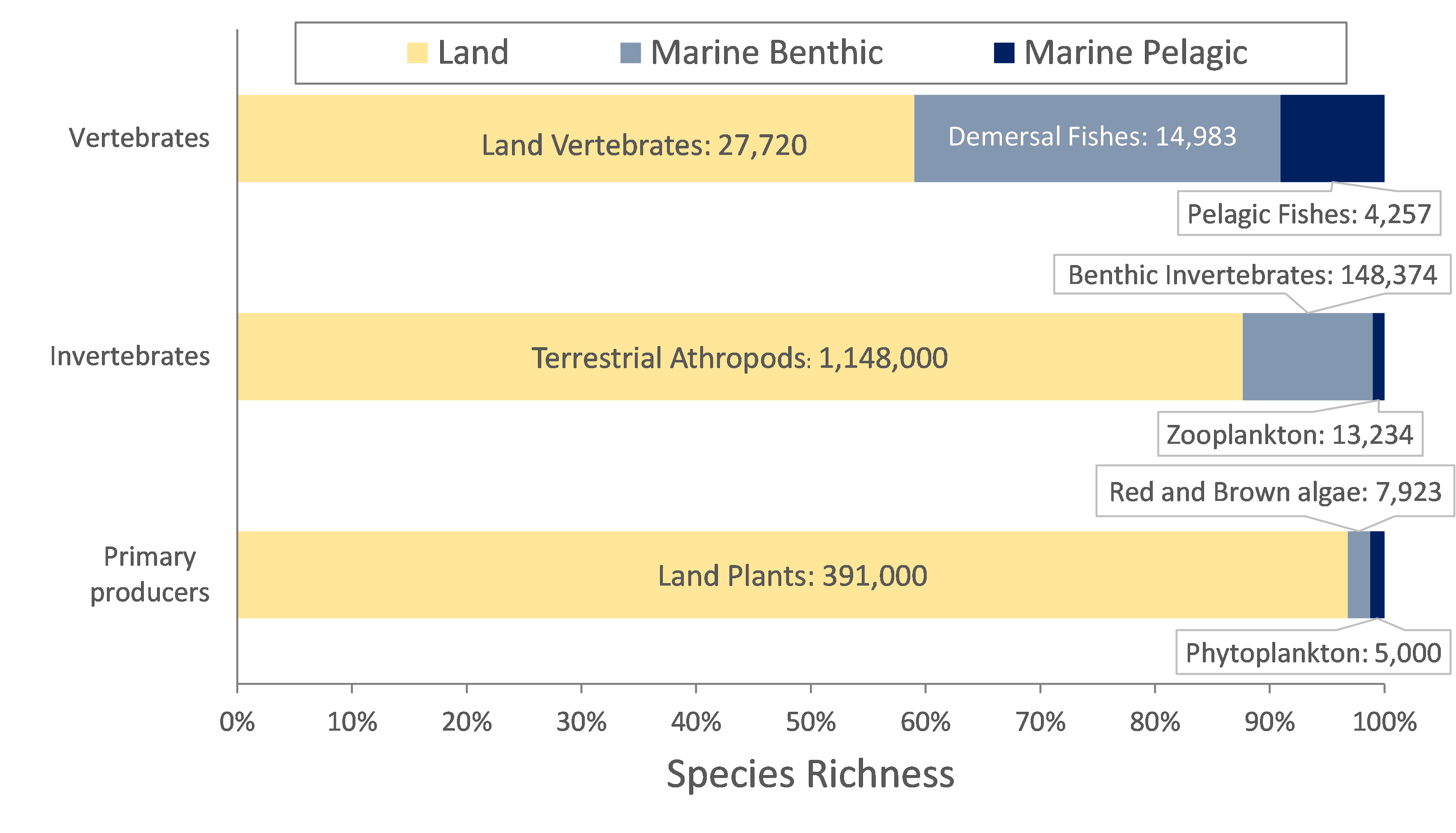
We reviewed the total species richness across taxa, trophic groups, and realms, according to the existing literature and a compilation of global data, shown in Fig. 1. Within low trophic levels, we found significant differences in diversity patterns between marine and land realms, particularly when comparing sunlit benthic and pelagic domains (Fig. 1). The species richness of primary producers in land and freshwater environments (391,000 species of vascular plants) is two orders of magnitude higher than that of marine eukaryotic phytoplankton (5,000 species). Diatoms are typically a dominant group in the pelagic domain, however, there are significantly more species in the benthic (8,770) relative to the pelagic domain (1,800). Arthropods diversified both in marine and land domains, and the described number of marine species is also substantially smaller than that of land species (Fig. 1). Concerning vertebrates, similar number of species has been described for fishes, as for terrestrial representatives (mammals, birds, reptiles, and amphibians combined). However, marine fishes are only half of the total fish diversity.
In summary, significant differences in species richness between marine and land realms are found particularly in primary producers and invertebrates, and mainly when comparing sunlit benthic from pelagic domains. The physical structure of the environment is therefore at the basis for understanding the disparity in diversity patterns.
At the global scale, the continental shelves include the areas with the highest regional species richness, such as the tropical coral reefs, in contrast to the pelagic domain.
Stokes’ law, drifting plankton, and the Sargasso Sea
The Ocean is in continuous motion, and so all seas are potentially physically connected, regardless of strong differences in oceanographic and chemical properties that generate biogeographic pelagic provinces. One of the key differences is the capacity of some organisms to attach to or live in close association with soil or the seafloor and maintain position, in contrast to the pelagic domain. The marine pelagic domain is a fluid environment characterized by near continuum horizontal and vertical axes where planktonic organisms drift passively by four main mechanisms: Brownian diffusion, sedimentation (i.e., sinking), convection, and advection. The sinking velocity of particles through the water column increases with the weight of the particle according to the Stokes’ law, hence shaping the body size of planktonic organisms (i.e., larger plankton cells tend to sink faster than smaller ones). This process, termed the Stokes’ law, mainly determines the small size of phytoplankton. In the open ocean, photosynthetic organisms must either afloat or be small enough that their sinking rates are slow and they stay in the light.
In addition to sinking, advection process is also suggested to be important in terms of body size in plankton. A recent study by Villarino et al. (2018) in Nature Communications on plankton β-diversity revealed a negative significant relationship between body size and dispersal scales for taxa ranging from bacteria to small fishes. This suggests that less abundant large-bodied communities have significantly shorter dispersal scales than more abundant small-bodied plankton. Larger planktonic organisms with longer generation times and smaller population densities might be more sensitive to stochastic local extinctions, loss of gene flow, and ecological drift. Consequently, we suggest that body size for passively dispersing organisms with sexual reproduction in open ocean is constrained not only by the abovementioned sinking process, but also by ocean advection, via the logarithmic decline between the geographic range of a species and its body size. We tested this hypothesis by relating the body size of 368 phytoplankton species and 412 zooplankton marine species with an estimate of species geographic range. The log-log relations are weak but significantly negative for both groups (r=-0.21 for phytoplankton, r=-0.22 for zooplankton, p<0.001), as expected by our hypothesis for plankton. Large free-floating primary producers are indeed extremely scarce in the ocean. The advection in the open ocean can easily isolate the individuals and propagules of a macroalgae species, making the population inviable due to longer generation time. Notably, the only macroalgae species with their entire lifecycle in the pelagic zone are the two species of Sargassum (S. natans and S. fluitans), that dwell in the Sargasso Sea, situated in the North Atlantic Subtropical Gyre where currents might facilitate the population cohesion of these macroalgae.
If a species can move to circumvent the water viscosity and to face the current advection (i.e., nekton), then, it might actively maintain the population cohesion. Thus, species´ geographic ranges might change according to their body size and dispersal mode (passive, active). For active dispersers, the negative relation is hence suggested to be lost or even reversed. We also tested this hypothesis by relating the body length of 215 marine pelagic species with an estimate of species geographic range. The relation is weak but significantly positive (r=0.14, p<0.036), supporting our hypothesis for active dispersers.
The theory of the limitations of pelagic diversity
We provide an integrative approach to explain why the marine realm is less diverse in terms of overall species richness than land, based on the primary effect of sinking and oceanic drift to body size of eukaryotic species with passive dispersal, which affects higher trophic levels:
Sinking and body size in marine primary producers. The key physical difference between pelagic ocean and sunlit benthos (and land) is the possibility for non-motile organisms (such as primary producers) to maintain position and hence population contiguity, as in land. In pelagic environments, primary producers, are small enough to avoid sinking due to Stokes’ Law and nutrient uptake efficiency. Large population densities and short generation times of planktonic organisms are needed to avoid extinction in open sea. In benthos, individuals are maintained in the system and eventual population collapse can recover under favourable conditions through latent phases. This would explain for instance why the diversification rate of benthic diatoms is higher than that of pelagic ones. As body size determine the mobility capacity and habitat complexity, it has in turn two different implications in the marine diversity.
Dispersal ability and population density in pelagic organisms. According to the mobility capacity of organisms and their density, advection and other processes are combined with sinking process to determine diversity patterns:
- Advection in high-density and widely distributed plankton populations. The higher connectivity of oceans, facilitated by advection, mix the plankton populations of the different seas, and hence may limit allopatric speciation in organisms with high density and wide distribution, compared with fragmented terrestrial habitats among islands and continents, or among river basins. This may explain why the rate of diversification of diatoms is higher in pelagic freshwaters than in marine, and why the number of marine free-living copepod species is not much higher than that of freshwater copepods as would be expected due to the different area of both habitats.
- Advection in low-density and restricted distribution plankton populations. Oceanic advection tends to isolate rare (i.e., very low density) passive organisms with restricted distribution that exceed a certain body size threshold and would reinforce the extinction process (via Stokes’ law). These processes may explain the absence of sexually reproducing macroalgae in open sea, while there are extant species of macroscopic free-living plants in freshwater.
- In nekton. Larger, active organisms can circumvent the oceanic drift and maintain mating encounter rates for population viability, which might explain that the richness of marine fishes is more balanced with that of terrestrial vertebrates than richness of arthropods and primary producers.
Habitat complexity and coevolution of primary producers in the pelagic domain. In contrast to macroalgae forests in the sunlit benthos, marine primary producers are small in open ocean and hence cannot generate habitat complexity, which limits the diversification of the upper trophic levels in the pelagic ecosystem. Small body size in plankton in turn constrains the diversification of functional traits, limiting the possibility of mutualist interactions such as facilitation, and hinders the emergence of habitat complexity by bioengineers, hence reducing niche differentiation and speciation, and limiting species coevolution. These processes may explain the higher overall richness and β-diversity of invertebrates in sunlit benthos with respect to zooplankton communities.
Isolation by distance across aquatic realms. Isolation and dispersal limitation among freshwater watersheds and among coastal fragments of sunlit benthic habitats promote higher allopatric speciation than in open oceans, which are more connected by currents and spatial configuration. These processes may explain the higher local and β-diversity fish richness in freshwaters and in coastal areas, such as the coral reef biodiversity hotspots, with respect to pelagic communities.
Area of the pelagic and sunlit benthic domains. Marine systems are dominated by pelagic ocean (63% of the surface area), which is significantly less locally diverse than sunlit benthos. Due to the relatively small global surface of sunlit benthos (8%) compared to land, the species-area relationship limits the overall marine species richness.
Acknowledgements
This study has been supported by the Horizon Europe research and innovation programme under grant agreement no. 101059915 (BIOcean5D project). Co-Funded by the European Union. Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union. Neither the European Union nor the granting authority can be held responsible for them.
The BRTA is a consortium that remains a step ahead of future socio-economic challenges worldwide and in the Basque Autonomous Community; it addresses them through research and technological development, thus projecting itself internationally. The BRTA centres collaborate to generate knowledge and transfer it to Basque society and industry so as to make them more innovative and competitive. The BRTA is an alliance of 17 R&D centres and cooperative research centres with the support of the Basque Government, the SPRI and the Chartered Provincial Councils of Araba, Bizkaia and Gipuzkoa.
References
- Chust, G., L. Alonso-Sáez, E. Villarino, J. Chave, N. Rodríguez-Ezpeleta, E. Arevalo, D. Righetti, and X. Irigoien (2025) Why do oceans shelter fewer species than land? A review on the role of sinking, dispersal and body size. Oikos doi: 10.1002/oik.11542 ↩