Life and deeds of RNA (I): William Wallace and the fate of the cell
Life and deeds of RNA (I): William Wallace and the fate of the cell

Among all the molecules that constitute living organisms, DNA (deoxyribonucleic acid) is probably the most popular and certainly it deserves the “molecule of life” surname. However, its role as the keeper of the critical information needed to construct an organism would be useless without the help of a very close relative, with a strikingly similar chemical composition but subtle, yet critical differences. We are talking about ribonucleic acid or RNA, and in the present series of posts we will revise some quite novel and fascinating molecular processes controlled exclusively by the RNA version of some genes.
DNA owes its fame to the fact that it encodes the information corresponding to proteins, in the form of discrete pieces of information that we call genes. For several decades the correspondence one gene = one protein and the role of RNA as an intermediate molecule used for of converting the information encoded into a set of four molecules (nucleotides) into the broader spectrum of twenty amino acids (which serve as the bricks used for construction of proteins) was assumed as a “central dogma of molecular biology” by Watson and Crick themselves. Unfortunate decision, calling a scientific theory “a dogma” (Francis Crick was later conscious of how wrong this concept was; being a genius does not save you to fail at some times). After some years and with the quick acquisition of genomic knowledge, this central idea soon was replaced by a dynamic view of the molecular machinery of the cell, where nucleic acids and proteins interact constantly in order to perform cellular functions. In this context, several other functions of RNA apart from its main role involving the transcriptional and translational processes, started to be described. In the last decades these novel findings gave rise to a large family of different RNA types regarding their size and function in the cell. And lastly, providing one of the most revolutionary molecular tools: RNA interference. But we will discuss this notion in the next article of the present series. Right now, it is time to find out what has William Wallace to do with RNA.
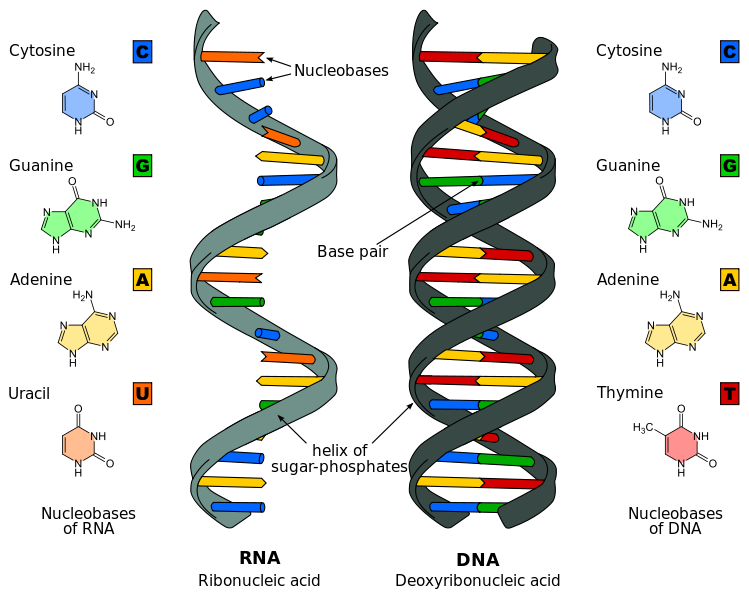
First of all, we should bear in mind that not all the information in the genome encodes proteins; there are long and long stretches of DNA which act as spacers between genes, or as targets for proteins which must bind specific regions promoting conformational changes that in turn promote or inhibit the expression of certain proteins. But before proteins, DNA creates a copy of itself in the form of RNA. And in some cases, certain genes are transcribed into RNA… and that’s all. There is not a corresponding protein. There are non-translated RNAs which vary in size and perform different tasks in the cell. The one we want to focus on belongs to the class of long, non-coding RNA or briefly, lncRNA. It may seem a novelty for the reader, but this molecules are known since years ago. Particularly a very well known example is Xist, a non-translated RNA involved in the mechanism of silencing the expression of an entire X chromosome in female mammals. This is a very interesting process that would deserve an entire article for itself, so let’s just keep the notion that a long chain of ribonucleic acid can be transcribed with the only mission of interacting with a particular molecular device, affecting its function.
In a multidisciplinary work, including computational, stem cell and developmental biologists and recently published in Cell 1, one of these lncRNAs and its critical function is described. Klattenhoff and collaborators were interested in the analysis of this kind of molecules in the area of cardiogenesis. Thanks to the increasingly settled technology of stem cells, researchers are able to manipulate populations of embryonic cells in order to study the critical processes that take part during organogenesis: in the present case, they analyzed the gene expression profile of mouse embryonic stem cells (ESCs) searching for the genes with the highest activation; then they compared with cells from adult tissues. Finally, they found a gene highly expressed in ESCs and also in heart. And thus we get to the Scottish hero, since they chose to call this gene Braveheart.
This molecular version of William Wallace turned out to be a chain of RNA formed by about 600 nucleotides, and they found no protein codified in its sequence. In a very ironic twist, the way to study this RNA was using another RNA against it. By using the technique of RNA interference, the researchers could study the effect of depleting Braveheart in ESCs: in doing so, they could learn not only in which way having or not Braveheart affected the cells, but also what other genes and the expression of which proteins was affected or not by its absence. Summarizing, the conclusions are that Braveheart is not a gene required for the survival of the embryonic cells; but is critical to make them transform into cardiac cells.
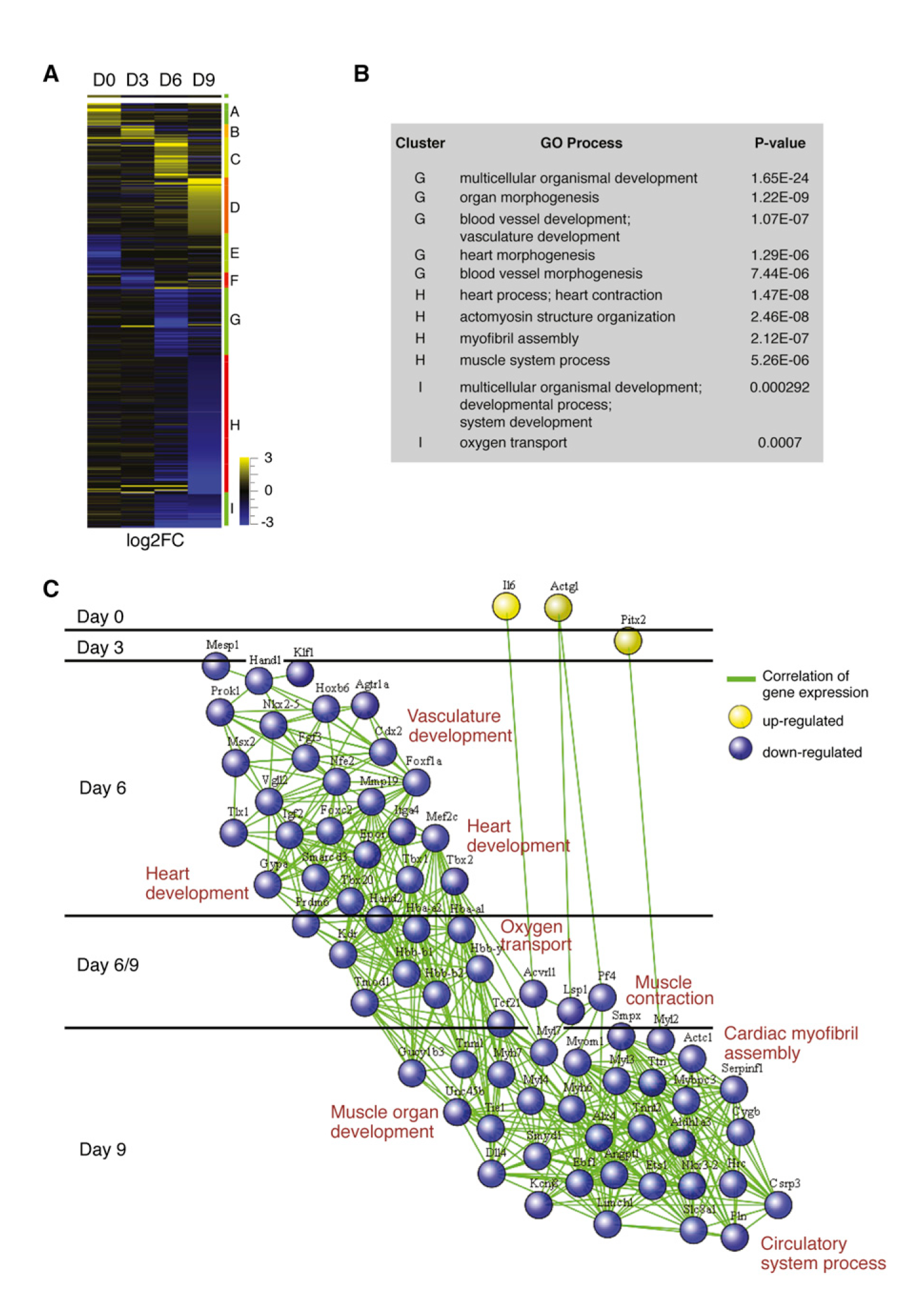
Let’s stop here to remember that the processes involved in the development of organisms require the initiation of genetic programs with a very precise timing of events: “packs of genes” are switched on and/or off in a concrete progression that guarantees the correct development of structures that assure the construction of an organism, cell by cell. Stem cell populations mutate and expand responding to these changes in genetic profiles, losing on their way the capacity to produce different lineages and commiting themselves to a particular one. The interesting finding here is how critical results the activity of this long, non translated RNA in determining that the cells in which it is present embrace the fate of becoming precursors of the cardiovascular system. And this is achieved by activating a critical switch, a protein called MESP1 that is able to activate a series of genes implicated in the development of all cell types in the heart. But how do Braveheart and MESP1 interact? They don’t. The effect of Braveheart, promoting the actions of MESP1, appears to function by an interaction mediated by other player called SUZ12. SUZ12 is a repressor, an enemy of Mesp1 and other genes that promote certain fates for a cell; when Braveheart attaches to SUZ12, impedes its repressive action and Mesp1 is free to start the revolution, making cells run towards a cardiac fate. Braveheart tasks do not stop here, since it is also critical for the correct physiology of cardiac cells in more advanced stages of differentiation. Not bad for a non-translated gene…
The relevance of this finding in a clinical context is obvious and encouraging: cardiovascular regeneration is a very limited process and surely not strong enough as to reverse severe injury after cardiovascular disease takes place; hence the knowledge of molecular determinants for cardiac cell populations and the techniques based on stem cells may constitute a turning point for the treatment of cardiovascular pathologies. There is yet a final difficulty: Braveheart has been found and described in the mouse genome, but there is not known equivalent in humans: the genes for lncRNAs appear to be highly species-specific, although their functions seem to be conserved and thus a human candidate hopefully exists, but remains to be unfold.
Probably the name chosen by these researchers was only due to the object of their study, a joke on cardiac cells; but curiously what they found shows striking parallelisms with the story of the revolutionary warrior portrayed in Mel Gibson’s film: a lonely warrior which promotes revolution of the cells, attacking the repressor that holds them away from their fate. Surely, it would be a piece of surprise for the warriors of ancient Scotland to know that their gestures form part of the discovery of cellular processes.
References
- Klattenhoff, C. A., Scheuermann, J. C., Surface, L. E., Bradley, R. K., Fields, P. A., Steinhauser, M. L., Ding, H., et al. (2013). Braveheart, a long noncoding RNA required for cardiovascular lineage commitment. Cell, 152(3), 570–83. doi:10.1016/j.cell.2013.01.003 ↩
2 comments
[…] the previous article of this series starred by ribonucleic acids, we found out about how an RNA molecule is able to regulate complex […]
I like the helpful information you provide in your articles.
I will bookmark your weblog and check again here frequently.
I am quite certain I will learn many new stuff right here!
Good luck for the next!