How do flowers release their perfume?
How do flowers release their perfume?

Plants emit a great amount of volatile organic compounds (VOCs). They are mainly produced in flowers and fruits and confer them their classical pleasant smell. However, leaves and roots also emit an important quantity of volatiles.
VOCs have different functions mostly related with the communication of the plant with the environment. They are responsible of making flowers attractive to pollinators and fruits to seed dispersers. Each plant species is capable of synthesizing a unique set of volatiles. Flowers generally emit the highest amount and most diverse mix of VOCs, which is species-specific and tends to reflect the type of pollinator, such as moths, bees or bats.
Besides, VOCs also protect plants from pathogens and parasites. For example, in vegetative organs, VOCs may be synthesized in surface glandular trichomes and be released due to mechanical disruption, such as after herbivore attacks as part of the plant defence machinery (see video above). Moreover, they also fulfil an important role in plant-plant communication since VOCs can act as allelochemicals and also as neighbouring detection signals, providing information about potential competitors. Some VOCS are also able to protect the plants against abiotic stresses, such as high light, temperature or oxidative stress. The overall importance of VOCs for plant performance is evident; indeed, it has been estimated that around 10% of the carbon fixed during photosynthesis is directed towards their synthesis.
In agriculture, VOCs are important to control crop-pollinator interactions and also for fruit consumer liking as reported for instance in my previous post about tomato flavour. VOCs are also essential for the industry of floriculture, flavour, cosmetic and fragrances.
VOCs are chemically diverse, at present close to 2000 different molecules have been identified but this list is continuously growing with the increase of the number of species being examined together with the development of more accurate analytical methods. They are divided into several classes including terpenoids, phenylpropanoids/benzenoids, fatty acid derivatives and amino acid derivatives. In spite of such chemodiversity, plant VOCs are generally low-molecular weight molecules, lipophilic and present high vapour pressure at ambient temperatures.
Because of those physicochemical properties, until recently it was accepted that VOCs could freely cross membranes and evaporate into the surrounding atmosphere. However, by computer simulation, it has been hypothesized that the emission of most VOCs by diffusion, linked to their lipophilic nature, would provoke their accumulation in cell membranes. That excessive accumulation would then disturb the proper functioning of membrane; therefore, suggesting the existence of active transport systems 1.

A recent work 2 led by Prof. Natalia Dudareva of Purdue University (Lafayette, Indiana, USA) tackled this question and investigated the existence of protein-mediated export of VOCs (Adebesin et al., 2017). To do so, the authors chose the flowers of petunia (Petunia hybrid; Figure 1) as working system. Indeed petunia is an important model in plant genetics and molecular biology, mostly used for the study of flower development, floral pigmentation and plant reproduction.
First, Adebesin et al. searched for ABC-type transporters that were highly expressed in petunia flowers. They chose these transporters because they are known to transport other lipophilic compounds such as diterpenes. ABC transporters constitute one of the biggest protein families. Generally, they are formed by two transmembrane domains that form a pore and two cytosolic domains that include an ATP-binding cassette that gives the name to the family (ABC) with ATPase function. With this approach Adebesin et al. found a gene of the ABC subfamily G, the gene PhABCG1 that was almost specifically expressed in petals of opened flowers.
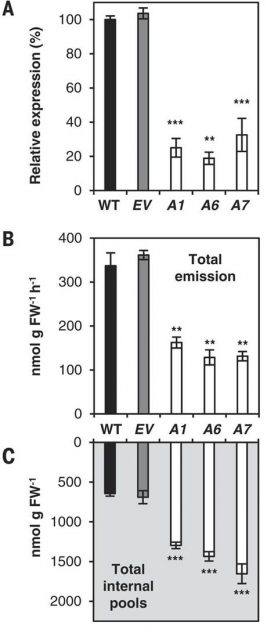
To test whether PhABCG1 transporter was involved in VOCs emission, the authors genetically transformed petunia to generate plants with reduced expression of this gene. To do so, they used RNA interference technology (RNAi) and selected three independent petunia lines that showed reduced PhABCG1 expression (around 70-80% reduction). Interestingly, these lines displayed decreased VOCs emission that was accompanied by the accumulation of internal VOCs (Figure 2).
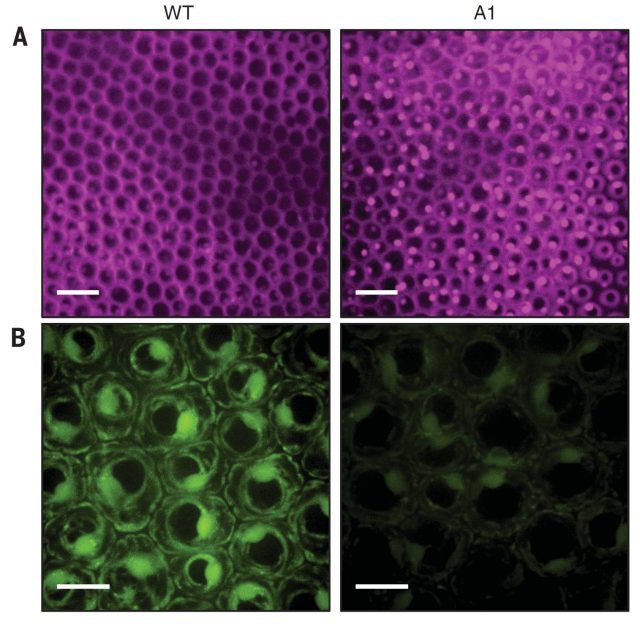
As mentioned above, it was hypothesized that VOCs accumulation could potentially have negative impact on membranes functionality. To test that hypothesis, the authors observed petunia flower cells by confocal microscopy and confirmed that the membrane integrity of RNAi transgenic petunia plants, that internally accumulated VOCs (Figure 2C), was disrupted (Figure 3).
In addition, Adebesin et al. supplied radiolabelled methylbenzoate and benzyl alcohol, two of the major petunia VOCs, to tobacco cells transformed with PhACBG1 gene and observed that these cells were actively transporting these two VOCs out of the cells. Thus, confirming the potential function of PhABCG1 in active VOCs transport (Adebesin et al., 2017).
Overall, the discovery that PhABCG1 is involved in active efflux of volatiles provides the first direct proof of a biological mechanism of VOCs emission and generates many questions. For instance, about the existence of other transporters, about the specificity of the transport towards different VOCs and related to how these transporters are able to modulate VOCs emission. Moreover, it opens a new gate for metabolic engineering of flowers perfume, and plant volatiles in general.
References
- Widhalm JR, Jaini R, Morgan JA and Dudareva N (2015) Rethinking how volatiles are released from plant cells. Trends in Plant Science. 20: 545–550. ↩
- Adebesin F, Widhalm JR, Boachon B, Lefèvre F, Pierman B, Lynch JH, Alam I, Junqueira B, Benke R, Ray S, Porter JA, Yanagisawa M, Wetzstein HY, Morgan JA, Boutry M, Schuurink RC and Dudareva N (2017) Emission of volatile organic compounds from petunia flowers is facilitated by an ABC transporter. Science 356: 1386–1388. ↩