Playing with the sex of melons and cucumbers
Playing with the sex of melons and cucumbers

Plants reproduce sexually to ensure genetic variability and thus, to continuously evolve in a constantly changing environment. In angiosperms, flowers are the sexual organs and most angiosperms are hermaphroditic, this means they have bisexual flowers with both male and female organs (Figure 1). Stamens are the male part of the flower, they are filaments ended by anthers where the pollen is produced. The female part of the plant is the pistil where the upper part is the stigma which receives the pollen and the bottom part is the ovary which contains the ovules. Fertilized ovules will generate the seeds which are commonly protected inside a fruit.
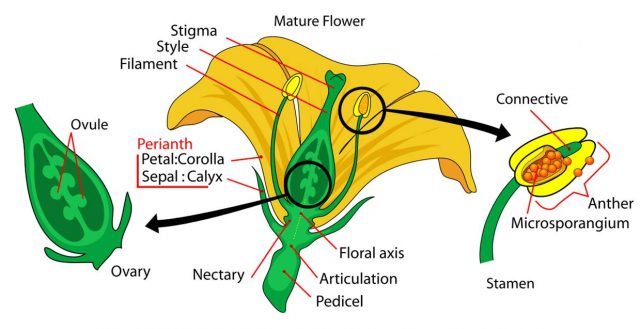
About ten percent of angiosperms have unisexual flowers. Approximately half of these species are monoecious, where male and female flowers coexist in the same individual plant, and the other half are dioecious species, where each plant individual only has male or female flowers. Among the species with unisexual flowers, we can find economically important crops such as the plant family Cucurbitaceae that includes watermelon, squash, cucumber or zucchini. Understanding flowering in general and sex determination of flowers, in particular, has important agronomic applications since flowers are at the origin of fruit formation. Thus, in the case of unisexual flowers, increasing the number of female flowers in monoecious species or the individuals with female flowers in dioecious species has great value to improve crops yield.
In consequence, the establishment of male and female traits has been extensively studied, mainly in hermaphroditic flowers and requires a genetic network that includes hormone regulators, microRNAS or transcription factors. Very recently (6th November 2015) a study of the research group of Prof. Abdelhafid Bendahmane, published in Science journal, provided a significant advance in the genetic control of sex determination in plants using as the models of study melons and cucumbers.
Previous studies of the same research group discovered two important genes for flowers sexual determination in monoecious melon, Cucumis melo (Boualem et al., 2008; Martin et al., 2009). The andromonoecious gene CmACS-7 which controls the development of stamina (male organs) in a female flower; indeed, when this gene is not functional female flowers become hermaphrodite. The gynoecious gene CmWIP1 whose expression leads to male flowers development; thus, its loss-of-function provokes purely female plants. CmACS-7 encodes 1-aminocyclopropane-1-carboxylic acid synthase (ACS), an enzyme for ethylene biosynthesis, an important hormone that regulates many plant processes. CmWIP1 encodes a C2H2 zinc finger transcription factor. This two genes work coordinately to control flowers sexual phenotype; CmWIP1 loss of function leads to CmACS-7 expression resulting in exclusive female flowers 1 2. In contrast, in those works, melon plants with exclusive male flowers were not described.
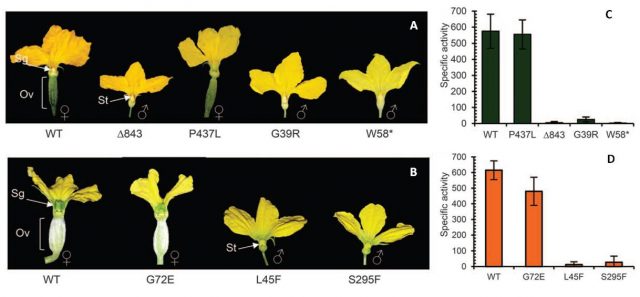
In this new study 3, the authors discovered the gene ACS11 that leads to androecy (exclusive presence of male flowers) in cucumber. ACS11 catalyzes the synthesis of the ethylene precursor ACC (1-aminocyclopropane-1-carboxylic acid). In Figure 2 it can be observed how different mutations in ACS11 sequence that compromised its activity affected the sex of the plant leading to androecy in both cucumber and melon.
Interestingly, the authors also observed that when CmACS11 was not expressed in melon the expression of CmWIP1 was induced suggesting a coordinated action of both genes. To understand their co-regulation, they studied a double mutant lacking both genes and those plants contrarily to ACS11 mutant did not lead to androecy but to a gynoecious phenotype (exclusive female flowers presence) therefore indicating that ACS11 acts upstream WIP1. Overall, the temporal and spatial control of ethylene production kinetics is crucial for flowers sex determination.
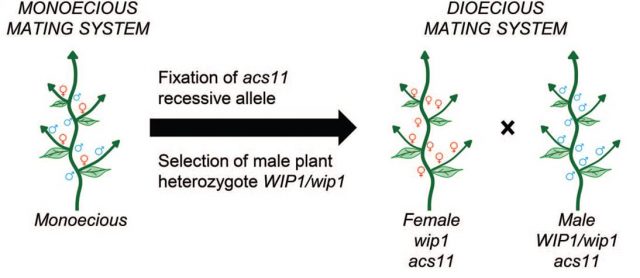
Finally, the authors were able to create artificial dioecious melon plants with a sex ratio 1:1 from a monoecious parental by generating plant populations with different combinations of acs11 and wip1 alleles (Figure 3). This works is a valuable step towards the understanding the control of flower development and may be of great importance to increase certain crops yield.
References
- Boualem A, Fergany M, Fernandez R, Troadec C, Martin A, Morin H, Sari MA, Collin F, Flowers JM, Pitrat M, Purugganan MD, Dogimont C, Bendahmane A (2008) A conserved mutation in an ethylene biosynthesis enzyme leads to andromonoecy in melons. Science. 2008 321:836-8. doi: 10.1126/science.1159023. ↩
- Martin A, Troadec C, Boualem A, Rajab M, Fernandez R, Morin H, Pitrat M, Dogimont C, Bendahmane A (2009) A transposon-induced epigenetic change leads to sex determination in melon. Nature 461:1135-8. doi: 10.1038/nature08498. ↩
- Boualem A, Troadec C, Camps C, Lemhemdi A, Morin H, Sari MA, Fraenkel-Zagouri R, Kovalski I, Dogimont C, Perl-Treves R, Bendahmane A (2015) A cucurbit androecy gene reveals how unisexual flowers develop and dioecy emerges. Science. 350:688-91. doi: 10.1126/science.aac8370. ↩
1 comment
I’m sure this research is important, but your writing style needs some work for readers to care. May I recommend reading The Forest Unseen by David Haskell for some insights on how to write well and clearly about plants?