The ape who learned to run: Were we born for endurance races?
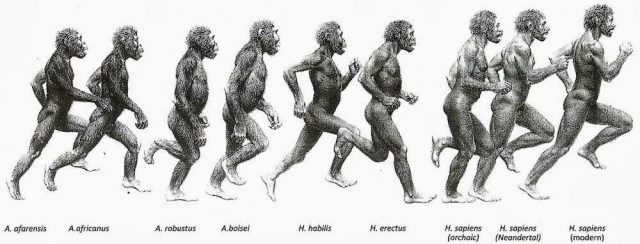
They say that we were born to run, not metaphorically as in that famous song by Bruce Springsteen, but more literally. The cursorial human being is neither the fastest nor the agilest or the most powerful (a simple backpack complicates things quite a lot). However humans, despite their quaint upright figure, present some breakneck numbers at endurance running (ER). Even without talking about elite athletes but about recreational runners. An average 20 to 40-year old person trained for endurance running is able to run a popular flat marathon, like the one in London in 2014, in an average time of 3 hours, 44 minutes (a The Telegraph report). Therefore, keeping 11km/h (3 m/s) along 42km. Long distance elite runners show stratospheric marks that an average person would barely beat even riding a mountain bike.
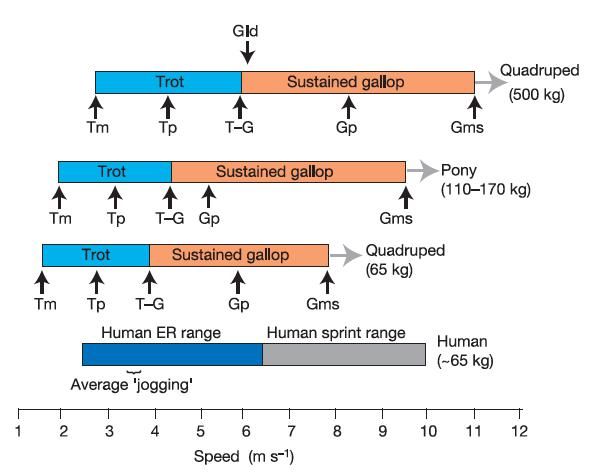
Aware of the numbers, our peculiar biped biology and some paleontological cues, The endurance running hypothesis began to take shape with a work of David R. Carrier in 19841. It attempts to explain several of our unique anatomical and physiological features from an evolutionary point of view as adaptations to the endurance running for hunting and scavenging. From that view, our ability to dissipate body heat by the lack of hair, the upright position and the ability to sweating (higher that most of the mammals); our invariable energy efficiency in a wide range of running speeds (consuming almost the same amount of energy from 8 to 20km/h); and our peerless respiratory cycle uncoupled from the gait (unlike the quadrupeds) would all be favoured by natural selection as valuable hunting tools. In fact, already the australopithecines were clearly bipeds and consumed some meat from medium-sized mammals lacking any known weapon. Therefore, exist the possibility that the early hominids could use persistence hunting or active fast scavenging for competing against other carnivores.
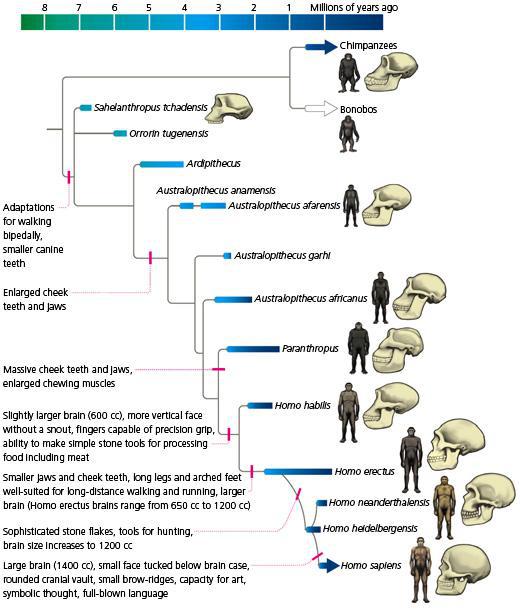
In persistence hunting, the prey is pursued from few hours to a couple of days until it collapses exhausted. Although almost extinct, such an art has been reported in several populations during the twenty-century. Bushmen, Tarahumara, Navajo or some Australian aborigines were skilled running down zebras, wildebeest, dears, or antelopes by ER; mainly during hot weather conditions. Those prey animals just to travel by walking, but they can trot as fast as a human run. They can also gallop faster than humans run but just for a while (Figure 2). However, their respiratory constrictions (breathing coupled with gait) and less efficient heat dissipation (often by panting) make them have preferred running speeds (Figure 2, Tp and Gp). In some species, those preferred speeds are below or about those that conditioned humans can sustain in ER. If the runners chasing a prey are able to track it, keeping good average speed for enough time under a burning sun, the preys will be pushed to run at their uneconomical gaits overheating their body’s until collapse.
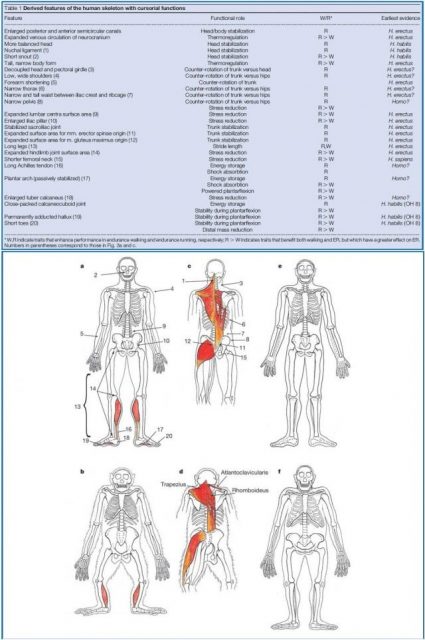
In addition to above considerations, a full set of biomechanical and musculoskeletal adaptations for running in genus Homo were exposed by Bramble and Lieberman in 2004, consolidating The Endurance Running Hypothesis in a joint body2. In this work, they reasoned a number of features of our body whose existence makes more evolutive sense for running than for walking. Furthermore, they also traced them back to its first evidence in the hominid line, in some cases even to the Homo habilis (Figure 3, upper panel). Some examples of these traits are long spring-like tendons (like Achilles tendon and iliotibial track), foot arch and relative reduction in distal limb mass; all involved in running economy. Working in running stabilization we find the gluteus maximus, the large erector spinal muscles and the nuchal ligament, which is also present in other cursorial mammals like horses. We also find comparatively larger articular surface areas in lower body joints for accommodating the stress generated by impact forces at running, which are, at least, twofold higher than in walking. Many of those features, while useful for running are little or not required for walking and most of them are undeveloped or absent in other primates (Figure 3, lower panel).
Therefore, the early hominids, lacking of any weapon, were less powerful and slower in peak speed than large predators and scavengers, which often act towards evening. Hence, the evolutionary pressure would have favoured the endurance running pursuing preys or looking for carcases during in the sweltering mornings. Making possible an increasing amount of protein in hominid diets during Pliocene and Pleistocene. Thus, helping in a dietary change assumed a key player in the development of the intelligence and behaviour of modern humans.
Besides and beyond the anatomy and bioenergetics, Kiely and Collins3 consider various processes in our neuromechanical system as adaptations for bipedal racing. In fact, our way of running on two legs seems, at first, terribly dangerous. However, the high impact loads at every step, the high energy demands and the lack of stability of such a “vertical” running, appear not to harm or fatigue our bodies as much as they should. The reason could hide in a superior running coordination. The cerebello-cerebral cortical network in Homo sapiens is relatively larger than in any other mammal. This structure seems to enable humans to generate models of his own movement with high spatial and temporal resolution. Helping to anticipate with extreme precision the requirements of the track and adapting the running dynamics for stability and efficiency. Playing, if necessary, to deal with more complex conditions or when fatigue diminishes the performance. Those models, acquired and refined with practice, modulate the rhythmic patterns generated by the Central Pattern Generators(CPG’s) in the spinal cord. The CPGs are able to keep the gait most of the time, leaving the brain free to other tasks like thinking and decision-making. Additionally, the same spine and brainstem generate the reflexes which modulate adaptation of movements to periodic disturbances. Finally, reflex movements, which are not guided by any neural instruction, would correct those little unforeseen imbalances occurred suddenly like with a misstep.
Nowadays, after millennia being farmers more dedicated to handling large weights or heavy armors, this hypothesis is difficult to accept. Even more taking into account that paleoecological evidence suggests a paleohabitat formed by savanna-woodlands, where tracking and running are difficult given the abundant vejetación and poor visibility. Lacking also evidences about whether cognitive abilities of the first hominids were enough for such a demanding activity. And without going into how they were able to stock up on water for such an effort4. However, unlike other primates, our bones, our muscles, our ligaments, our cardiorespiratory system and our neuromechanical control have the mark of animals born not only for long walks. A man running a marathon spends the same energy whether he jogs or he runs at elite pace, and only 30% more than if he walks those 42km (Lieberman 2007). In the very beginning, a blending of group endurance hunting and a prey selection towards young or weak individuals would facilitate such a formidable task.
Evolution by natural selection does what it can with what it has. Therefore, despite our extravagant way of running among the animals, we are exceptionally good at it as a result of our innate versatility to face different terrains and long distances, in different weather conditions and at a remarkable pace. Rivaling in endurance to some species of medium-sized mammals. Apart few breeds of camels, horses and dogs, all established by men for endurance running, not many wild terrestrial animals are able to travel by foot the distances that many humans are able to run.
References
- Carrier, D. R. et al. The Energetic Paradox of Human Running and Hominid Evolution (and Comments and Reply) Current Anthropology Volume 25, Number 4, Aug. – Oct., 1984. ↩
- Bramble, D. M. & Lieberman, D. E. (2004) Endurance running and the evolution of Homo. Nature 432, 345–352. doi: 10.1038/nature03052 ↩
- Kiely, J. & Collins, D. J. (2016) Uniqueness of Human Running Coordination : The Integration of Modern and Ancient Evolutionary Innovations. Front. Psychol., doi: 10.3389/fpsyg.2016.00262. ↩
- Rayne, T. & Bunn, H. T. (2007)The endurance running hypothesis and hunting and scavenging in savanna-woodlands. J Hum Evol. 53, 434–438. ↩
2 comments
[…] Cada vez está de moda, no solo correr, sino correr maratones. Pero, ¿está el cuerpo humano, ese bípedo bajado de los árboles, preparado para ello? Daniel Moreno en The ape who learned to run: Were we born for endurance […]
[…] Image: mappingignorance […]