Metacognition in nonhumans
Metacognition in nonhumans
Metacognition is «cognition about cognition», or «knowing about knowing». It comes from the root word «meta», meaning beyond, and the word «cognition» that includes all mental abilities and processes related to knowledge: attention, memory, judgment and evaluation, reasoning, biological computation, problem-solving, decision making, comprehension and production of language, etc.
Metacognition can take many forms; it includes knowledge about when and how to use particular strategies for learning or for problem-solving and the evaluation of the own cognitive processes. There has been an intense debate in the scientific literature in recent years over whether metacognition is unique to humans.

A study in chimpanzees used a task that required them to use symbols on a keyboard to name what food was hidden in a location 1. The chimpanzees could either report the identity of the food immediately or first check a container in which the food had been hidden. The chimpanzees were significantly more likely to visit the container first on trials in which they could not know its contents but were more likely to just name the food item without looking into the container on trials in which they had seen its contents. This acting reflects a capacity to evaluate information that serves to support intelligent responding, and it strongly suggests that our closest living relative has metacognitive abilities closely related to those of humans 2.
But what about other more distant species such as social insects? Sometimes ants are seen as brainless machines, biological automata that keep doing the same processes and tasks even if everything around is altered. This is simply not true. For their ecological success, animals must continually gather information about their surroundings and respond appropriately to changes in this environment. One good example is foragers. These animals need some cognitive abilities such as a good memory and well-developed communicative skills informing their partners of specific aspects about food sources such as patch productivity, safety, and quality. These communication strategies use different types of pheromones including guide molecules that form a trail that goes from the food to the nest, therefore recruiting new foragers and increasing gathering.
If the system is not flexible, a changing environment can be a social catastrophe, sending workers where nothing can be found as pheromone trails cannot be quickly removed. Honeybees Apis mellifera can prevent nest-mates from recruiting to a dangerous food source by using a stereotyped ‘stop’ signals and Pharaoh’s ants, Monomorium pharaonis, are reported to deposit a ‘no entry’ pheromone marker on the trail that does not lead to a food source. However, there is no information for other ants but it seems that the no entry signal is not generalized.
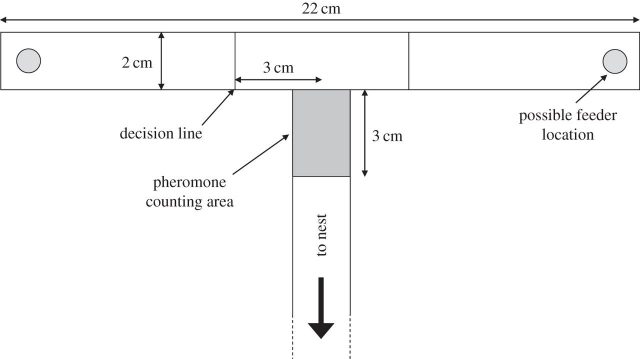
Another strategy has been recently observed in ants 3. It consists of «overwriting» obsolete information upregulating the production of new information, i.e. upregulating pheromone trail deposition in response to changes in the location of food. The authors, Tomer J. Czaczkes and Jürgen Heinze, trained ants to a feeder location, and then altered the environment by changing the feeder location to the other arm of a T-maze. Then, the ants first do not found food where it previously was and after wandering around, they find food but in a different location. In the ant society, collective decision-making systems are characterized by positive feedback: one worker finds some food, she recruits more workers to the food, and they in turn recruit more workers. This allows colonies to make rapid and accurate collective decisions. However, such positive-feedback systems move rapidly towards local optima, where they can become trapped. For example, if an ant colony finds a poor food source and recruits heavily to it, it should not be able to switch to a newly discovered high-quality food source, because the first pheromone trail is too strong.
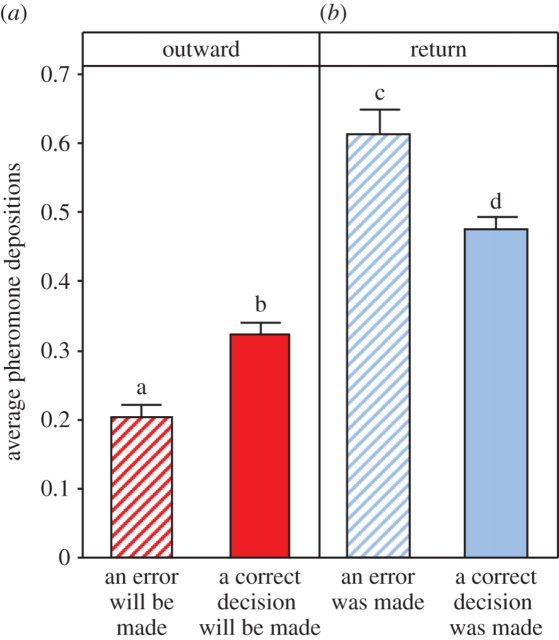
Trained ants that had found a changed feeder position dramatically increased the number of pheromone depositions on their way back to the colony. This was true of both ants which had correctly followed their memory to the old (now empty) feeder location, and those ants which had mistakenly taken the wrong path to the new (now rewarding) feeder location. Then, ants responded to an environmental change by strongly up-regulating pheromone deposition immediately after experiencing the change. This may help maintain the colony’s foraging flexibility, and allow multiple food locations to be exploited simultaneously. The treatment also caused uncertainty in the foragers, by making their memories less reliable. Ants returning to the nest after eventually finding the food source up-regulated pheromone deposition if they had made a wrong choice. An alternative behavior, which does not require a memory, would be for ants to upregulate pheromone when returning on an unmarked path, and in fact, pheromone deposition of returning ants is higher on unmarked paths.
The apparent response of ants to making an error by upregulating pheromone deposition on the return journey has been described previously. Intriguingly, the authors also found that outgoing ants which went on to make an error deposited less pheromone. This seems to imply that the ants can measure the reliability of their own memories, and respond accordingly: if the ant does not trust its own memory, it deposits less pheromone. The ability to assess the quality of a memory (known as metamemory judgment, which is an aspect of metacognition) is a highly advanced cognitive ability.

It has been also investigated whether the honeybee possesses the capacity for metacognition in solving a complex visual discrimination task 4. Free flying bees were trained to enter a tube and choose a variable target above or below a black bar. Changing the angle of the target shape to the horizontal of the reference bar varied the difficulty of this task. Bees performed better on easier trials and worse on more difficult trials. Subsequently, bees were allowed to opt out of the current trial (by flying through an exit port) and begin a new trial. a subset of bees display behavior that is supportive of metacognition: 1) opt out more often as difficulty of task increases, 2) transfer the concept of opting out to a newly learned task, 3) perform better on trials where they have the option to avoid the decision and 4) take more time to opt out of more difficult trials. These results support the idea that bees have also some basic level of metacognitive ability in certain tasks. Metacognition, therefore, is not restricted to humans and can be accomplished in a much smaller and limited brain.
References
- Beran MJ, Smith JD, Perdue BM (2013) Language-trained chimpanzees (Pan troglodytes) name what they have seen but look first at what they have not seen. Psychol Sci 24(5): 660-666. ↩
- Metacognition: Ability to ‘think about thinking’ not limited to humans. Phys Org 3 April 2013 ↩
- Czaczkes TJ, Heinze J (2015) Ants adjust their pheromone deposition to a changing environment and their probability of making errors. Proc Biol Sci 282(1810). DOI: 10.1098/rspb.2015.0679 ↩
- Perry CJ, Barron AB (2013) Honey bees selectively avoid difficult choices. Proc Natl Acad Sci U S A 110(47): 19155–19159. ↩
1 comment
[…] Tú conoces. Además tu conoces que conoces. A esto es a lo que se llama metacognición, a saber que se sabe. Los humanos, la inmensa mayoría al menos, parece ser que metaconocemos. ¿Y los animales no humanos?¿Metaconocen?¿Metaconocen los chimpancés?¿Y […]